Research Article |
|
Corresponding author: Gretchen M. Walters ( gretchen.walters@unil.ch ) Academic editor: Brecht Verstraete
© 2022 Gretchen M. Walters, Diosdado Nguema, Raoul Niangadouma.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Walters GM, Nguema D, Niangadouma R (2022) Flora and fire in an old-growth Central African forest-savanna mosaic: a checklist of the Parc National des Plateaux Batéké (Gabon). Plant Ecology and Evolution 155(2): 189-206. https://doi.org/10.5091/plecevo.85954
|
Abstract
Background and aims – Old-growth savannas in Africa are impacted by fire, have endemic and geoxylic suffrutices, and are understudied. This paper explores the Parc National des Plateaux Batéké (PNPB) in Gabon and the impact of fire on its flora to understand if it is an old-growth savanna. It presents 1) a vascular plant checklist, including endemic species and geoxylic suffrutices and 2) an analysis of the impact of fire on the savanna herbaceous flora, followed by recommendations for fire management to promote plant diversity.
Material and methods – 1,914 botanical collections from 2001–2019 collected by the authors and others were extracted from two herbaria databases in 2021 to create the checklist. The impact of fire was explored through a three season plot-based inventory of plant species (notably forbs and geoxylic suffrutices) in five annually, dry-season burned study areas located at 600 m in elevation. A two-factor ANOVA was conducted across two burn treatments and three season treatments.
Key results – The area has a vascular flora of 615 taxa. Seven species are endemic to the Plateaux Batéké forest-savanna mosaic. Seventeen species are fire-dependent geoxylic suffrutices, attesting to the ancient origins of these savannas. Burning promotes fire-dependent species.
Conclusion – The PNPB aims to create a culturally-adapted fire management plan. The combination of customary fire and fire-adapted species in the savanna creates a unique ancient forest-savanna mosaic in Central Africa that merits protection while recognising the role that the Batéké-Alima people have in shaping and governing this landscape.
Keywords
checklist, Plateaux Batéké, Gabon, fire, forest-savanna mosaic, floristics, Central Africa, national park, ancient grassy biome, old growth savanna, geoxylic suffrutices
Introduction
In recent years, research on the importance of savanna ecosystems has shown that while major conservation and restoration efforts have focused on forested ecosystems, savannas in some parts of the world have remained understudied and their biodiversity value relatively unknown or undervalued (
However, recent research on African savannas provides evidence of their ancient origins (
In Central Africa, savanna ecosystems and their floral diversity in relation to fire remain underexplored; studies focus on carbon stocks and biomass (
The Plateaux Batéké are covered with a forest-savanna mosaic situated within the Guineo-Congolian centre of endemism (


The Plateaux Batéké landscape. A. Customary fire in the savanna near Kele la Tsiere. B. Kele la Kalami (Mont Kalami), a sacred site and the highest point in the study site at 700 m. C. Riparian forest. D. Abandoned village forests within the savanna. E. Cirque. F. Mopia Bai, from the major forest block. All photos were taken by Gretchen Walters.
How old are these savannas? In general, climate fluctuation between arid and moist periods favoured savanna and forest expansion at different times: an arid period favoured savanna formation 40,000–70,000 years BP, followed by a wet period of “forest revival” around 30,000 years BP, followed by an arid period until 18,000 years BP where savanna resurged, followed by a wetter period 3000 years BP (
Although our study does not aim to date the study site savannas of the Plateaux Batéké, other studies have made several estimations from different sites across the plateaus. In the colonial era, the Plateaux Batéké savannas, like elsewhere in Africa, were thought to be of anthropogenic origin (
No similar studies have been conducted in the Gabonese portion of the plateaus including in the study area and therefore it cannot be confirmed if similar vegetation responses to climate are found there. However, a alternative way to understand if these savannas are ancient is through their vegetation composition. Ancient and old-growth savannas comprise species communities, which require centuries to assemble (
Across Gabon, the savannas are of different ages and typically of climatic origin, originating during cooler periods when savannas were expanding. While those of the Plateaux Batéké are estimated to have first formed 30,000 to 70,000 years BP (
Beyond climate, fire is an important factor shaping savannas. Anthropogenic savanna fires in the Plateaux Batéké have occurred since at least 2,100 years BP (
Although fire and vegetation studies have been the object of many long-term experiments in Africa, dating from colonial times (
Material and methods
The study area comprises the PNPB and a buffer zone of 25 km. This wider zone was included for two reasons: 1) to include botanical collections that had been made in similar habitats, but outside the park, and 2) for the fire study to have access to areas that were regularly burned and accessible to compare with the single area from the park which had not burned for three years. This area was located close to park camps, discouraging the presence of hunters that typically light fires elsewhere in the park, and so the area remained unburned at the time of the study.
Checklist
Botanical specimens have been collected from the PNPB area since 2001 by a variety of collectors, and they are most notably deposited in LBV, MO, and WAG (herbarium acronyms follow Thiers continuously updated). Although the area was traversed by several Europeans during the colonial era, who provided vegetation descriptions and descriptions of fire use in the 1800s (
The majority of plant collections were identified at BR, LBV, MO, and the National Herbarium of the Netherlands (NHN), and registered in the databases at LBV, MO, and WAG. Database records were obtained in 2016 from the MO and WAG databases and again from MO in 2021 for the area comprising PNPB and including a 25 km limit of the Park. There were no specimens from the Republic of Congo in the circumscribed area. This represented 1,921 collections that have been collected in all seasons, from 2001 to 2019, in part due to residential collection (by the first author) from 2006 to 2008.
Endemic species were determined by their classification in the Checklist of Gabon (
Collection density was analysed by importing all collection coordinates, where reported, into QGIS v.3.16.4 (
Fire and flora
A subset of the savanna flora from the checklist was studied to understand the impact of fire on forb diversity and geoxylic suffrutices, based on the 183 savanna species from a previous study derived from the study area data set (
In each area, five hilltop sites at approximately 600 m in elevation were selected and revisited each season for one year. There is variation in sand type by stratigraphy (
Species numbers from the three subplots were grouped together into a single cumulative plot; this became the unit for comparison. Each treatment (burned, unburned) was visited in the long-dry (July 2007), mid-rainy (December 2007), and late-rainy (April 2008) seasons producing two sets of data (burned and unburned sites) across three seasons. Thus, for each of the three seasons, 30 plots could be compared; for each burn treatment, 45 plots could be compared. A total of 90 plots were available for comparison (Table
| Season | Total plots per burn treatment | ||||
| Long-dry | Mid-rainy | Late-rainy | |||
| Burn treatment | Burned plots | 15 | 15 | 15 | 45 |
| Unburned plots | 15 | 15 | 15 | 45 | |
| Totals plots per season | 30 | 30 | 30 | 90 | |
Normality of the distribution of species per treatment was tested by calculating the mean and standard deviation per treatment and then determining if 70% of the values fell within this range (
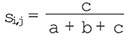
Where c is the number of species in common, a is the number of species only occurring in treatment i and b is the number of species only occurring in treatment j (
Results
Checklist
The checklist is the first published checklist of the PNPB and comprises 612 species (615 taxa), representing 105 families (Supplementary file 1). Fabaceae, Rubiaceae, and Poaceae are the top three families, with the top ten families representing 55% of all species (Table
| Family | Number of taxa |
| Fabaceae | 83 |
| Rubiaceae | 77 |
| Poaceae | 40 |
| Cyperaceae | 30 |
| Apocynaceae | 24 |
| Annonaceae | 20 |
| Asteraceae | 17 |
| Ochnaceae | 17 |
| Euphorbiaceae | 14 |
| Melastomataceae | 13 |
| Genus | Number of taxa |
| Cyperus | 13 |
| Psychotria | 12 |
| Campylospermum | 10 |
| Vigna | 9 |
| Landolphia | 7 |
| Dalbergia | 6 |
| Vernonia | 6 |
| Utricularia | 6 |
| Millettia | 6 |
Four species of the checklist have mostly Congolian distributions but reach the edge of their western-most distribution in the gallery forests of the Plateaux Batéké, as confirmed by recent collections: Dracaena waltersiae (
Of the 1,914 collections from the data set, 1,897 were able to be mapped, with coordinates. Of these, 70% have been collected within the PNPB, and largely in the northern half, with significant collections around the following localities of the park: the “Débarcadère” in the north, Lac Loulou to the east, the Project Protection des Gorilles main camp and Mbié Cirque in the centre, and Bai Djobo in the western forest. The focus on the northern part of the PNPB is due to the difficulty in access: the park is largely bounded by rivers with no bridges, lacks roads, and must be crossed by foot, quad, or boat. The remaining 30% of the collections were made in the proposed area for PNPB to be extended, including around Ekouyi-Mboma village, due to the resident collection by the first author in that area. Collections also include the Kele la Kalami rock outcrop, Lewou Cirque, and the forb-fire study area (see Fig.

Fire, forbs and geoxylic suffrutex diversity
The forbs and geoxylic suffrutices in this part of the study represent a subset of savanna species in the checklist: 49 hilltop species (see Supplementary file 2), comprising 28% of the overall savanna flora, based on the 183 savanna species from a previous study derived from the study area data set (
2-Way ANOVA results showing that only season was a significant factor in affecting species diversity. An asterisk “*” denotes significance.
| Source | Type III SS | d.f. | Mean Squares | F-ratio | p value |
| BURN | 6.533 | 1 | 6.533 | 2.378 | 0.124 |
| SEASON | 34.956 | 2 | 17.478 | 6.361 | 0.002* |
| BURN x SEASON | 8.956 | 2 | 4.478 | 1.630 | 0.198 |
| Error | 725.422 | 264 | 2.748 |
When Tukey’s HSD was used to determine which seasons were most significant, the dry season differed significantly from both rainy season in terms of species numbers, while the rainy seasons were not significantly different from one another (Table
Tukey’s HSD determine which seasons were significantly different from each other in terms of species numbers. Here, the p-values are displayed. An asterisk “*” denotes significance.
| Dry | Mid-Rainy | Late-Rainy | |
| Dry | 0.000 | – | – |
| Mid-Rainy | 0.036* | 0.000 | – |
| Late-Rainy | 0.002* | 0.584 | 0.000 |
To understand if burning affects species diversity within the burn treatments, species lists from the 90 plots (see Supplementary file 2) of burn treatments were compared using the Jaccard Similarity Index. The dry season is the most divergent, having only 32% species in common with the other seasons (but 45% for mid-rainy season and 50% for the late-rainy season). However, when species lists were pooled by season alone, the dry and late-rainy season were the most dissimilar, with 36% in common (for 44% for dry-mid rainy; 50% for mid- and late-rainy). Overall, whether considering species or season, the species overlap was always 50% or less.
However, it is insufficient to consider only species numbers and seasons. To understand if some species were only found in the burned or unburned plots, across the 90 plots, species were compared by burn treatment and season: 17 species are found only in burned areas, representing 35% of the species in the fire study, while three occur only in unburned areas. The dry season has the highest number of unique species, while the late- and the mid-rainy seasons comprise fewer. In order to understand how fire affects these 17 species, the botanical and ecological literature for each species was consulted to understand if the species exhibits Fire Stimulated Flowering (FSF) (Table
Species unique to burned plots, with observations on fire-related aspects of species biology.
| Family | Species | Observations on fire-dependent species |
| Apocynaceae | Glossostelma lisianthoides | The genus Glossostelma has “erect stems coming up after fires from narrow, tuberous, perennial rootstocks…” ( |
| Apocynaceae | Xysmalobium holubii | Genus noted to comprise “pyrophytic herbs of fire-prone grasslands” ( |
| Asparagaceae | Dipcadi viride | Bulb ( |
| Asteraceae | Helichrysum mechowianum var. ceres | Perennial herb. Species noted to be in “regularly burned areas” ( |
| Asteraceae | Vernonia daphnifolia | Perennial herb with wood rootstock. After fires, the rootstock grows numerous herbaceous stems ( |
| Asteraceae | Vernonia guineensis | Perennial herb with woody rootstock. Found in burned savannas being the “premier recur” ( |
| Asteraceae | Vernonia potamophila | Geoxylic suffrutex ( |
| Caryophyllaceae | Polycarpaea eriantha | Although found in fire-prone habitats, information related to fire was not found. |
| Fabaceae | Eriosema glomeratum | Geoxylic suffrutex. Flowering after fire ( |
| Fabaceae | Eriosema pellegrini | Geophytic herb ( |
| Fabaceae | Eriosema shirense | Herb with woody rootstock ( |
| Fabaceae | Macrotyloma biflorum | Perennial herb ( |
| Fabaceae | Vigna oblongifolia var. oblongifolia | Annual herb ( |
| Hypoxidaceae | Curculigo pilosa | Rhizome. Favoured by fire ( |
| Lamiaceae | Kalaharia schaijesii | Geoxylic suffrutex ( |
| Orobanchaceae | Buchnera paucidentata | Annual herb, hemiparasitic ( |
| Orobanchaceae | Striga asiatica | Annual herb, hemiparasite ( |
The results from this part of the study show that while season heavily influences savanna forb diversity, fire is also important in the life cycle of many forbs and geoxylic suffrutices, and that savanna burning promotes forb species.
Discussion
Floristic diversity
This study shows that the floristic diversity, across 615 species, comprises seven endemic species and 17 geoxylic suffrutices, as well as four Congolian species which reach their limits in the study area. Furthermore, the area, though represented by 1,914 collections, shows a collection bias for the northern part of the PNPB.
Previous analysis of the area’s flora showed both a strong element of Lower Guinea species (
Eriosema batekense and Kalaharia schaijesii have been described in the past decade or so (
New floristic inventories of the proposed park extension (see next section) have recently begun and it is expected that new species and records will be discovered, especially if new areas are visited. This study presents a baseline from which to work. Future botanical work should focus on the forest, especially its riverine forests, but also the under-collected areas of the southern part of the PNPB. Our checklist shows few forest flora endemic species, however, Memecylon batekeanum (
The interior savannas of Gabon, such as those of PNPB, continue to be explored, with other floristic studies underway around Franceville and Moanda. Those areas are expected to be floristically distinct from the PNBP. For example, from a Haut Ogooué checklist of ferns, largely collected around Franceville, few of the 84 species reported were found in the PNPB (
Given the age of first savanna formation in the Plateaux Batéké of 30,000–70,000 years ago, and a persistence of savanna in many places in the Plateaux area over time, it is not surprising that they contain endemic species and geoxylic suffrutices. However, beyond climatic origins of the savanna habitat and associated flora, anthropogenic factors also contribute to the floristic diversity: people have created village forests or copses, and initiate fire regimes. The Batéké-Alima have long created copses in open savanna (
Fire and floristic diversity
In this study, beyond maintaining an open, savanna environment and related diversity, fire appears to be important for floristic diversity in two ways: fire fosters the emergence of some species post-fire and it favours some life forms such as geoxylic suffrutices.
Emergence of forbs in burned areas
Our study shows little connection between forb diversity and fire, when explored by species numbers by burn treatments and season. However, even though fire typically stimulates flowering within two years (
Early studies from the Plateaux Batéké area described the relationship of fire to the flora.
However, the annual fire regimes at the time of these observations were disrupted during the colonial and post-colonial periods. In the 1960s, just after Gabon’s independence from France, fire regimes changed from annual, dry season fires, to semi-annual fires, with these latter being more frequent, cooler fires, which can be classified as a “frequent, cool, small” regime, which occur on an annual or semi-annual basis (
In studies in other grasslands in Africa, fire does not consistently affect forb diversity. In South Africa, fire regimes had no effect on forb diversity, although there are distinct fire-tolerant and fire-intolerant floras (
Presence of geoxylic suffrutices
We report seventeen species as geoxylic suffrutices. The presence of this life form is an indicator that the savannas of the PNPB are old-growth, showing a link between the presence of fire in an ecosystem and its adapted flora (
Fire management
In Africa, fire is currently used to achieve a variety of park management goals (
Maps of worldwide forest restoration opportunities have misclassified this area (the entire Plateaux Batéké) as one that should be targeted for tree planting (
In order to maintain a diversity of plant species, including high forb diversity and species that are fire-dependent, a patch mosaic approach (
Potential extension of the PNPB
In terms of protection, the area under consideration for the extension would extend the PNPB by adding 344,460 acres north of the park. This proposed area contains some areas considered to be sacred including Mopia Bai and Kele la Kalami (Mont Kalami), a rock outcrop. The rocky outcrops of the area occur at higher elevations in the savanna, typically at 600–700 m. Their vegetation is distinct, along with unique bird species (e.g. the African River Martin, Pseudochelidon eurystomina) and plant species populations including Polystachya dendrobiiflora and Bidens oligoflora. The area is under customary management with restricted access and constitutes a type of protection similar to a Territory of Life (
Conclusion
The savannas of the PNPB are old-growth savannas as demonstrated by their age, the presence of endemic savanna species, and a fire-dependent flora which includes geoxylic suffrutices. The PNPB aims to develop a culturally-adapted fire management plan; this study provides evidence that burning favours some species which flower post fire, or whose life form depends on fire. The combination of customary fire and land traditions which have created unique habitats, and the presence of fire-adapted savanna species create a cultural landscape in Gabon that merits continued protection while recognising how the Batéké-Alima have shaped and continue to govern this landscape.
Acknowledgements
Plant identifications were contributed by Roy Gereau, Marc Sosef, Jos van der Maesen, Steven Dessein, Olivier Lachenaud, Cornelius E.N. Ewango, David Harris, Sylvia Phillips, David Goyder, Petra de Block, Elmar Robbrecht, Jan Wieringa, Frans Breteler, David Kenfack, and Carel Jongkind. Identifications by GW, RN, and DN were carried out at BR, K, LBV, MO, and WAG. Rainfall records for 2006 were provided by the Projet Protection des Gorilles. The authors especially thank Liz Pearson, Paul Aczel, and Sandra Mahé for their assistance with organising fieldwork within the PNPB. We also thank the residents of Ekouyi-Mboma for welcoming GW and many other biologists and social scientists to stay in their village. Egid Onas, Djo Kewemie, George Kandinia, Prince Bissiemou, Etienne Mounoumoulossi, John Stone, Adam Bradley helped conduct fieldwork and collect plants. Marina Cracco assisted with the mapping of collection density and Raphaël Bubloz, Marina Cracco, and Vasco Ferreira da Costa generated the maps. Jan Wieringa and Nicolas Texier provided database extracts of specimens. Library access to consult the botanical literature was provided by the Geneva Botanical Garden. Other literature was consulted at the Archives d’Outre-Mer (Aix-en-Provence, France). We thank Brecht Verstraete for his help in finalising the photo plate. We thank Brecht Verstraete, Elmar Robbrecht, and the anonymous reviewers for their comments that improved the manuscript.
References
- Aleman JC, Blarquez O, Elenga H, Paillard J, Kimpuni V, Itoua G, Issele G, Staver AC (2019) Palaeo-trajectories of forest savannization in the southern Congo. Biology Letters 15: 20190284. https://doi.org/10.1098/rsbl.2019.0284
- Aleman JC, Fayolle A, Favier C, Staver AC, Dexter KG, Ryan CM, Azihou AF, Bauman D, te Beest M, Chidumayo EN, Comiskey JA, Cromsigt JPGM, Dessard H, Doucet J-L, Finckh M, Gillet J-F, Gourlet-Fleury S, Hempson GP, Holdo RM, Kirunda B, Kouame FN, Mahy G, Gonçalves FMP, McNicol I, Quintano PN, Plumptre AJ, Pritchard RC, Revermann R, Schmitt CB, Swemmer AM, Talila H, Woollen E, Swaine MD (2020) Floristic evidence for alternative biome states in tropical Africa. Proceedings of the National Academy of Sciences 117: 28183–28190. https://doi.org/10.1073/pnas.2011515117
- ANPN (2008) Plan de Gestion du Parc National des Plateaux Batéké 2008–2013.
- Archibald S, Lehmann CER, Gómez-Dans JL, Bradstock RA (2013) Defining pyromes and global syndromes of fire regimes. Proceedings of the National Academy of Sciences 110: 6442–6447. https://doi.org/10.1073/pnas.1211466110
- Aubame J-H (1947) Renaissance Gabonaise: Programme de Regroupement des Villages. Imprimerie Officielle, Brazzaville.
- Aubréville A (1949) Climats, forêts et désertification de l’Afrique tropicale. Société d’Editions Géographiques, Maritimes et Coloniales, Paris.
- Aubréville A (1953) Les expériences de reconstitution de la savane boisée en Côte d’Ivoire. Bois et Forêts des Tropiques 32: 4–10.
- Aubréville A (1962) Savanisation tropicale et glaciations quaternaires. Adansonia série II 1(16): 16–84.
- Bamps P (2013) A new species of Kalaharia (Lamiaceae) from Central Africa. Plant Ecology and Evolution 146: 134–137. https://doi.org/10.5091/plecevo.2013.746
- Barnett DT, Stohlgren TJ (2003) A nested-intensity design for surveying plant diversity. Biodiversity and Conservation 12: 255–278. https://doi.org/10.1023/A:1021939010065
- Batsa Mouwembe MA, Ifo SA, Binsangou Malonga S, Koubouana F (2017) Variabilité spatiale des stocks de carbone organique du sol des savanes dans les réserves de Lésio-Louna et de Léfini, plateaux Tékés, République du Congo. Afrique Science 13(3): 297–307.
- Baudon A (1929) Contribution à l’étude des plantes oléagineuses de l’Afrique Equatoriale. Annales du Musée colonial de Marseille série IV 7(12): 27.
- Beentje H (2000) New taxa and new combinations in Helichrysum (Compositae: Inuleae). Kew Bulletin 55: 349. https://doi.org/10.2307/4115647
- Beentje HJ (2021) Flore du Gabon. Volume 56: Compositae. Margraf, Weikersheim.
- Biaou SSH (2009) Tree recruitment in west African dry woodlands: the interactive effects of climate, soil, fire and grazing. PhD thesis, Wageningen University, the Netherlands.
- Bond WJ (2016) Ancient grasslands at risk. Science 351: 120–122. https://doi.org/10.1126/science.aad5132
- Bond WJ, Parr CL (2010) Beyond the forest edge: ecology, diversity and conservation of the grassy biomes. Biological Conservation 143: 2395–2404. https://doi.org/10.1016/j.biocon.2009.12.012
- Bond W, Zaloumis NP (2016) The deforestation story: testing for anthropogenic origins of Africa’s flammable grassy biomes. Philosophical Transactions of the Royal Society B: Biological Sciences 371: 20150170. https://doi.org/10.1098/rstb.2015.0170
- de Brazza PS (1887) Voyages dans l’Ouest Africain par Monsieur Savorgnan de Brazza. 1875–1887. Textes et dessins inédits. Tour du Monde: 289–336.
- Bremond L, Oslisly R, Sebag D, Bentaleb I, Favier C, Henga-Botsikabobe K, Mvoubou M, Ngomanda A, de Saulieu G, Ecotrop Team (2021) Establishment and functioning of the savanna marshes of the Lopé National Park in Gabon since the termination of the African humid period and the arrival of humans 2500 years ago. The Holocene 31: 1186–1196. https://doi.org/10.1177/09596836211003230
- Brockett BH, Biggs HC, van Wilgen BW (2001) A patch mosaic burning system for conservation areas in southern African savannas. International Journal of Wildland Fire 10: 169. https://doi.org/10.1071/WF01024
- Brookman-Amissah J, Hall JB, Swaine MD, Attakorah JY (1980) A re-assessment of a fire protection experiment in north-eastern Ghana savanna. The Journal of Applied Ecology 17: 85. https://doi.org/10.2307/2402965
- Cardoso AW, Oliveras I, Abernethy KA, Jeffery KJ, Lehmann D, Edzang Ndong J, McGregor I, Belcher CM, Bond WJ, Malhi YS (2018) Grass species flammability, not biomass, drives changes in fire behavior at tropical forest-savanna transitions. Frontiers in Forests and Global Change 1: 6. https://doi.org/10.3389/ffgc.2018.00006
- de Chavannes C (1935) Avec Brazza: Souvenirs de la Mission de l’Ouest-Africain (mars 1883–1886). Plon, Paris.
- Christy P (2008) Le Feu et les Oiseaux de Savane. Internal Report to Batéké Plateaux National Park, Lékoni, Gabon.
- Clist B (1995) Gabon: 100,000 Ans d’Histoire. Centre Culturel Français, Libreville.
- Collin A (1951) Mise en défense contre les feux de brousse et reforestation des savanes du Bas-Congo. In: Conférence Forestière Interafricaine, Abidjan, décembre 1951, 441–458.
- Coquery-Vidrovitch C (1972) Le Congo au Temps des Grandes Compagnies Concessionnaires 1898–1930. Tome 1. Mouton & Co., Paris. https://doi.org/10.4000/books.editionsehess.359
- Cornelis D, Le Bel S, Mikolasek O, Caron A, Aubert S, Karpe P, Müller J-P (2017) Vers une gestion territorialisée de la chasse villageoise et des systèmes alimentaires carnés en Afrique centrale. In: Van Vliet N, Nguinguiri JC, Cornelis D, Le Bel S (Eds) Communautés Locales et Utilisation Durable de la Faune en Afrique Centrale. FAO, CIFOR, CIRAD, 143–174.
- Couvreur TLP (2008) Revealing the secrets of African Annonaceae: systematics, evolution and biogeography of the syncarpous genera Isolona and Monodora. PhD thesis, Wageningen University, the Netherlands. https://edepot.wur.nl/16321 [accessed 12.04.2022]
- Damen THJ, van der Burg WJ, Wiland-Szymańska J, Sosef MSM (2018) Taxonomic novelties in African Dracaena (Dracaenaceae). Blumea - Biodiversity, Evolution and Biogeography of Plants 63: 31–53. https://doi.org/10.3767/blumea.2018.63.01.05
- Dauby G, Leal M, Stévart T (2008) Vascular plant checklist of the Pongara National Park. Systematics and Geography of Plants 78: 155–216. https://www.jstor.org/stable/20649761 [accessed 12.04.2022]
- Dechamps R, Lanfranchi R, Le Cocq A, Schwartz D (1988) Reconstitution d’environnements quaternaires par l’étude de macrorestes végétaux. (Pays Batéké, R.P. du Congo). Palaeogeography, Palaeoclimatology, Palaeoecology 66: 33–44. https://doi.org/10.1016/0031-0182(88)90079-X
- Deklerck V, De Mil T, Ilondea BA, Nsenga L, De Caluwé C, Van den Bulcke J, Van Acker J, Beeckman H, Hubau W (2019) Rate of forest recovery after fire exclusion on anthropogenic savannas in the Democratic Republic of Congo. Biological Conservation 233: 118–130. https://doi.org/10.1016/j.biocon.2019.02.027
- Delègue M-A, Fuhr M, Schwartz D, Mariotti A, Nasi R (2001) Recent origin of a large part of the forest cover in the Gabon coastal area based on stable carbon isotope data. Oecologia 129: 106–113. https://doi.org/10.1007/s004420100696
- Demichelis C, Oszwald J, Bostvironois A, Gasquet-Blanchard C, Narat V, Bokika J-C, Giles-Vernick T (2021) A century of village mobilities and landscape dynamics in a forest-savannah mosaic, Democratic Republic of Congo. Bois & Forêts des Tropiques 348: 3–16. https://doi.org/10.19182/bft2021.348.a31934
- Dudley N (2008) Guidelines for Applying Protected Area Management Categories. International Union for the Conservation of Nature, Gland. https://doi.org/10.2305/IUCN.CH.2008.PAPS.2.en
- Dudley N, Jonas H, Nelson F, Parrish J, Pyhälä A, Stolton S, Watson JEM (2018) The essential role of other effective area-based conservation measures in achieving big bold conservation targets. Global Ecology and Conservation 15: e00424. https://doi.org/10.1016/j.gecco.2018.e00424
- Dupré M-C, Pinçon B (1997) Métallurgie et Politique en Afrique Centrale: Deux Mille Ans de Vestiges sur les Plateaux Batéké Gabon, Congo, Zaïre. Editions Kathala, Paris.
- Duvigneaud P (1949) Les savanes du Bas-Congo. Essai de phytosociologie topographique. Lejeunia, Mémoires 10: 1–192.
- Ebouli JM (2001) Les structures de type féodal en Afrique Centrale. Le cas des Téké: étude des relations de dépendance personnelle et des rapports de production entre ‘A mfumu’ et ‘Elogo dja Mfumu’ (des Origines à 1880). Master’s thesis, Université Omar Bongo, Gabon.
- Elenga H, Vincens A (1990) Paléoenvironnements quaternaires récents des Plateaux Batéké (Congo): étude palynologique des dépôts de la dépression du bois de Bilanko. In: Lanfranchi R, Schwartz D (Eds) Paysages Quaternaires de l’Afrique Centrale Atlantique. ORSTOM, Paris, 271–282. https://www.documentation.ird.fr/hor/fdi:34792 [accessed 12.04.2022]
- Elenga H, Schwartz D, Vincens A (1994) Pollen evidence of late Quaternary vegetation and inferred climate changes in Congo. Palaeogeography, Palaeoclimatology, Palaeoecology 109: 345–356. https://doi.org/10.1016/0031-0182(94)90184-8
- Fagan ME (2020) A lesson unlearned? Underestimating tree cover in drylands biases global restoration maps. Global Change Biology 26: 4679–4690. https://doi.org/10.1111/gcb.15187
- Fairhead J, Leach M (1996) Misreading the African Landscape: Society and Ecology in a Forest-Savanna Mosaic. First edition. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9781139164023
- Fayolle A, Swaine MD, Aleman J, Azihou AF, Bauman D, te Beest M, Chidumayo EN, Cromsigt JPGM, Dessard H, Finckh M, Gonçalves FMP, Gillet J-F, Gorel A, Hick A, Holdo R, Kirunda B, Mahy G, McNicol I, Ryan CM, Revermann R, Plumptre A, Pritchard R, Nieto-Quintano P, Schmitt CB, Seghieri J, Swemmer A, Talila H, Woollen E (2019) A sharp floristic discontinuity revealed by the biogeographic regionalization of African savannas. Journal of Biogeography 46: 454–465. https://doi.org/10.1111/jbi.13475
- Finckh M, Revermann R, Aidar MPM (2016) Climate refugees going underground – a response to Maurin et al . (2014). New Phytologist 209: 904–909. https://doi.org/10.1111/nph.13567
- Finckh M, Wendefeuer J, Meller P (2021) Frost‐driven lower treelines in Angola and their implications for tropical forest–grassland mosaics. Journal of Vegetation Science 32: e13084. https://doi.org/10.1111/jvs.13084
- Fischer E, Ghazanfar SA (2016) Schrophulariaceae s.l. In: Flore du Gabon 48, Margraf, Weikersheim, 89–149.
- Flügel TJ, Eckardt FD, Cotterill FPD (2015) The present day drainage patterns of the Congo river system and their neogene evolution. In: de Wit MJ, Guillocheau F, de Wit MCJ (Eds) Geology and Resource Potential of the Congo Basin. Springer, Berlin, Heidelberg, 315–337. https://doi.org/10.1007/978-3-642-29482-2_15
- de Foresta H (1990) Origine et évolution des savanes intramayombiennes (R.P. du Congo). II. Apports de la botanique forestière. In: Lanfranchi R, Schwartz D (Eds) Paysages Quaternaires de l’Afrique Centrale Atlantique. Editions de l’ORSTOM, Paris, 326–335.
- Fowler J, Cohen L, Jarvis P (1998) Practical Statistics for Field Biology. Second edition. John Wiley and Sons, Chichester.
- Fuhlendorf SD, Engle DM (2004) Application of the fire-grazing interaction to restore a shifting mosaic on tallgrass prairie: shifting mosaic on tallgrass prairie. Journal of Applied Ecology 41: 604–614. https://doi.org/10.1111/j.0021-8901.2004.00937.x
- Fynn RWS, Morris CD, Edwards TJ (2005) Long‐term compositional responses of a South African mesic grassland to burning and mowing. Applied Vegetation Science 8: 5–12. https://doi.org/10.1111/j.1654-109X.2005.tb00623.x
- Gami N (2003) Mission d’Information et d’Etude Socio-Economique dans les Villages de la Sous-Préfecture de Lékana (Congo Brazzaville), Frontalière du Parc National des Plateaux Batéké (Gabon). Rapport pour le Projet Protection des Gorilles (Gabon).
- Giresse P (1978) Le contrôle climatique de la sédimentation marine et continentale en afrique centale atlantique à la fin du quaternaire - problémes de corrélation. Palaeogeography, Palaeoclimatology, Palaeoecology 23: 57–77. https://doi.org/10.1016/0031-0182(78)90082-2
- Giresse P (2005) Mesozoic–Cenozoic history of the Congo Basin. Journal of African Earth Sciences 43: 301–315. https://doi.org/10.1016/j.jafrearsci.2005.07.009
- Giresse P, Kouyoumontzakis G (1990) Témoins quaternaires du littoral du Congo et du sud du Gabon. In: Lanfranchi R, Schwartz D (Eds) Paysages Quaternaires de l’Afrique Centrale Atlantique. ORSTOM Editions, Paris, 106–109. https://www.documentation.ird.fr/hor/fdi:34775 [accessed 12.04.2022]
- Giresse P, Maley J, Chepstow-Lusty A (2020) Understanding the 2500 yr BP rainforest crisis in West and Central Africa in the framework of the Late Holocene: Pluridisciplinary analysis and multi-archive reconstruction. Global and Planetary Change 192: 103257. https://doi.org/10.1016/j.gloplacha.2020.103257
- Gotelli NJ, Ellison AM (2004) A Primer of Ecological Statistics. Sinauer Associates, Inc., Sunderland, Massachusetts.
- Goyder DJ (1995) Notes on the African Genus Glossostelma (Asclepiadaceae). Kew Bulletin 50: 527. https://doi.org/10.2307/4110325
- Goyder DJ (2008) Xysmalobium samoritourei (Apocynaceae: Asclepiadoideae), a new species from the Guinea Highlands of West Africa. Kew Bulletin 63: 473–475. https://doi.org/10.1007/s12225-008-9059-1
- Goyder DJ (2009) A synopsis of Asclepias (Apocynaceae: Asclepiadoideae) in tropical Africa. Kew Bulletin 64: 369–399. https://doi.org/10.1007/s12225-009-9133-3
- Goyder DJ, Gonçalves FMP (2019) The Flora of Angola: collectors, richness and endemism. In: Huntley BJ, Russo V, Lages F, Ferrand N (Eds) Biodiversity of Angola. Springer International Publishing, Cham, 79–96. https://doi.org/10.1007/978-3-030-03083-4_5
- Guillot B (1980) La création et la destruction des bosquets Koukouya, symboles d’une civilisation et de son déclin. Cahiers O.R.S.T.O.M., série Sciences Humaines XVII(3–4): 177–189.
- Guiral L (1889) Le Congo Français. Du Gabon à Brazzaville. Plon, Paris.
- Gurney GG, Darling ES, Ahmadia GN, Agostini VN, Ban NC, Blythe J, Claudet J, Epstein G, Estradivari, Himes-Cornell A, Jonas HD, Armitage D, Campbell SJ, Cox C, Friedman Whitney R, Gill D, Lestari P, Mangubhai S, McLeod E, Muthiga NA, Naggea J, Ranaivoson R, Wenger A, Yulianto I, Jupiter SD (2021) Biodiversity needs every tool in the box: use OECMs. Nature 595: 646–649. https://doi.org/10.1038/d41586-021-02041-4
- Haddon IG (2000) Kalahari group sediments in the Cenezoic in southern Africa. Oxford Monographs on Geology and Geophysics, Oxford.
- Harris D, Armstrong KE, Walters GM, Wilks C, Mouandza Mbembo J-C, Niangadouma R, Wieringa JJ, Breteler FJ (2012) Phytogeographical analysis and checklist of the vascular plants of Loango National Park, Gabon. Plant Ecology and Evolution 145: 242–257. https://doi.org/10.5091/plecevo.2012.641
- He T, Lamont BB, Pausas JG (2019) Fire as a key driver of Earth’s biodiversity. Biological Reviews 94: 1983–2010. https://doi.org/10.1111/brv.12544
- Humphrey GJ, Gillson L, Ziervogel G (2020) How changing fire management policies affect fire seasonality and livelihoods. Ambio 50: 475–491. https://doi.org/10.1007/s13280-020-01351-7
- ICCA Consortium (2021) Territories of Life: 2021 Report. ICCA Consortium. https://report.territoriesoflife.org/ [accessed 21.04.2022]
- Ifo AS, Gomat HY, Mampouya Wenina YE, Lokegna DL, Minkala Nzonzi OR, Apendi Ngala GC, Henry M, Boundzanga GC, Jourdain C, Picard N, Loumeto JJ (2018) Carbon stocks and tree allometries in the savannahs of the Plateau Batéké, central Africa. Forest Ecology and Management 427: 86–95. https://doi.org/10.1016/j.foreco.2018.05.065
- IUCN (2011) A World of Opportunity for Forest and Landscape Restoration. International Union for the Conservation of Nature, Gland, Switzerland.
- Jacobs MJ, Schloeder CA (2002) Fire frequency and species associations in perennial grasslands of south‐west Ethiopia. African Journal of Ecology 40: 1–9. https://doi.org/10.1046/j.0141-6707.2001.00347.x
- Jeffery KJ, Korte L, Palla F, Walters G, White LJT, Abernethy KA (2014) Fire management in a changing landscape: a case study from Lopé National Park, Gabon. PARKS 20: 39–52. https://doi.org/10.2305/IUCN.CH.2014.PARKS-20-1.KJJ.en
- Jongkind CCH (2021) A review of Combretum falcatum (Welw. ex Hiern) Jongkind (Combretaceae) and related species from Africa, including Combretum baldwinii Jongkind, sp. nov., from Nigeria. Adansonia 43: 241–249. https://doi.org/10.5252/adansonia2021v43a22
- Keay RWJ (1959) Derived savanna – derived from what? Bulletin de l’Institut Français d’Afrique Noire, série A 21(2): 427–438.
- Koechlin J (1957) Morphoscopie des sables et végétation dans la région de Brazzaville. Bulletin de l’Institut d’Etudes Centrafricaines (COG) 13/14: 39–48.
- Koechlin J (1961) La Végétation des Savanes dans le Sud de la République du Congo (capitale Brazzaville). Mémoires ORSTOM, Imprimerie Charité, Montpellier. https://www.documentation.ird.fr/hor/fdi:15020 [accessed 12.04.2022]
- Kull CA (2004) Isle of Fire: the Political Ecology of Landscape Burning in Madagascar. University of Chicago Press, Chicago.
- Lamont BB, Downes KS (2011) Fire-stimulated flowering among resprouters and geophytes in Australia and South Africa. Plant Ecology 212: 2111–2125. https://doi.org/10.1007/s11258-011-9987-y
- Lamont BB, He T, Pausas JG (2017) African geoxyles evolved in response to fire; frost came later. Evolutionary Ecology 31: 603–617. https://doi.org/10.1007/s10682-017-9905-4
- Laris P (2004) Grounding environmental narratives: the impact of a century of fighting against fire in Mali. In: Moseley WG (Ed.) African environment and development. King’s SOAS Studies in Developmental Geography, London, 63–85.
- Laris P, Wardell DA (2006) Good, bad or “necessary evil”? Reinterpreting the colonial burning experiments in the savanna landscapes of West Africa. The Geographical Journal 172: 271–290. https://doi.org/10.1111/j.1475-4959.2006.00215.x
- Lebamba J, Ngomanda A, Vincens A, Jolly D, Favier C, Elenga H, Bentaleb I (2009) Central African biomes and forest succession stages derived from modern pollen data and plant functional types. Climate of the Past 5: 403–429. https://doi.org/10.5194/cp-5-403-2009
- Leeuwenberg AJM (ed.) (1985) Series of Revisions of Apocynaceae. Agricultural University Wageningen Papers.
- Lotte AJ (1953) Situation démographique du District de Franceville. Bulletin de l’Institut d’Etudes Centrafricaines 6: 161–180.
- Louppe D, Oattara N, Coulibaly A (1995) The effects of brush fires on vegetation. The Aubréville Fire Plots after 60 years. Commonwealth Forestry Review 74: 288–291. https://agritrop.cirad.fr/391053/ [accessed 12.04.2022]
- van der Maesen J, Walters GM (2011) Novitates gabonenses 77: A new Eriosema (Leguminosae-Papilionoideae) from Gabon and adjacent Congo. Plant Ecology and Evolution 144: 101–105. https://doi.org/10.5091/plecevo.2011.494
- van der Maesen LJG, Sosef MSM (2016) Flore du Gabon: Leguminosae-Papilionoideae. Margraf, Weikersheim.
- Maley J, Doumenge C, Giresse P, Mahé G, Philippon N, Hubau W, Lokonda MO, Tshibamba JM, Chepstow-Lusty A (2018) Late Holocene forest contraction and fragmentation in central Africa. Quaternary Research 89: 43–59. https://doi.org/10.1017/qua.2017.97
- Mangenot G (1955) Etudes sur les forêts des plaines et plateaux de la Côte d’Ivoire. Etudes Eburnéennes 4: 5–61.
- Maurin O, Davies TJ, Burrows JE, Daru BH, Yessoufou K, Muasya AM, Bank M, Bond WJ (2014) Savanna fire and the origins of the ‘underground forests’ of Africa. New Phytologist 204: 201–214. https://doi.org/10.1111/nph.12936
- Menaut J-C (1983) The vegetation of African savannas. In: Bourlière F (Ed.) Tropical savannas. Elsevier Scientific Publishing Company, Amsterdam, 109–149.
- Mpounza M, Samba-Kimbata MJ (1990) Aperçu sur le climat de l’Afrique centrale occidentale. In: Lanfranchi R, Schwartz D (Eds) Paysages quaternaires de l’Afrique Centrale Atlantique. Editions de l’ORSTOM, Paris, 31–41. https://www.documentation.ird.fr/hor/fdi:34766 [accessed 12.04.2022]
- Mundy NI (2000) New pteridophyte records from Gabon, West Africa with a preliminary species list for Haut-Ogooué Province. Fern Gazette 16(3): 125–142.
- Nieman WA, Van Wilgen BW, Leslie AJ (2021) A review of fire management practices in African savanna-protected areas. Koedoe 63. https://doi.org/10.4102/koedoe.v63i1.1655
- Nieto‐Quintano P, Mitchard ETA, Odende R, Batsa Mouwembe MA, Rayden T, Ryan CM (2018) The mesic savannas of the Bateke Plateau: carbon stocks and floristic composition. Biotropica 50: 868–880. https://doi.org/10.1111/btp.12606
- Nordal I, Iversen JI (1986) Hypoxidaceae. In: Flore du Gabon 28, Muséum national d’Histoire naturelle, Paris, 45–53.
- Obone C, Sosef MS (2010) Hyacinthaceae. In: Flore du Gabon 40, Margraf, Weikersheim, 19–30.
- Papy L (1949) Les Populations Batéké (A.E.F.). Cahiers d’Outre-Mer II (6): 112–134. https://www.persee.fr/doc/caoum_0373-5834_1949_num_2_6_1619 [accessed 12.04.2022]
- Parr CL, Brockett BH (1999) Patch-mosaic burning: a new paradigm for savanna fire management in protected areas? Koedoe 42: 117–130. https://doi.org/10.4102/koedoe.v42i2.237
- Parr CL, Andersen AN (2006) Patch mosaic burning for biodiversity conservation: a critique of the pyrodiversity paradigm. Conservation Biology 20: 1610–1619. https://doi.org/10.1111/j.1523-1739.2006.00492.x
- Pellegrin F, Le Testu G (1938) La Flore du Mayombe, Tome 3. Mémoires de la Société Linnéenne de Normandie, Caen.
- Perriguey N (1951) La conférence forestière inter-africaine d’Abidjan. Bois et Forêts des Tropiques 22: 96–103. https://revues.cirad.fr/index.php/BFT/article/view/18473 [accessed 12.04.2022]
- Peyrot B (1991) La géologie de l’Afrique Centrale. In: Lanfranchi R, Clist B (Eds) Aux origine de l’Afrique Centrale. Centre Culturel Français d’Afrique Centrale, Centre International des Civilisations Bantu, Libreville, 7–9.
- Phillips S (2016) Eriocaulaceae. In: Flore du Gabon 48, Margraf, Weikersheim, 1–20.
- Pigafetta P, Lopes D (1883) Le Congo: la Véridique Description du Royaume Africain, Appelé, Tant par les Indigènes que par les Portugais, le Congo, Telle qu’elle a Été Tirée Récemment des Explorations d’Edouard Lopez. Translated by Léon Cahun. J-J. Gay, Brussels.
- QGIS Development Team (2021) QGIS Geographic Information System. Version 3.16.4. http://qgis.osgeo.org [accessed 21.04.2022]
- Quammen D (2003) Saving Africa’s eden. National Geographic 203(9): 50–74.
- Rabinowitz D, Cairns S, Dillon T (1986) Seven forms of rarity and their frequency in the Flora of the British Isles. In: Soulé ME (Ed.) Conservation Biology: the Science of Scarcity and Diversity. Sinauer Associates, Inc., Sunderland, 182–204.
- Ramsay JM, Rose-Innes R (1963) Some quantitative observations on the effects of fire on the Guinea savanna vegetation of northern Ghana over a period of 11 years. African Soils 8: 41–120.
- Raynal J (1968) Itinéraires et lieux de récolte de Georges le Testu. In: Flore du Gabon 14, Muséum National d’Histoire Naturelle, Paris, 17–66.
- Revermann R, Gonçalves FM, Gomes AL, Finckh M (2017) Woody species of the Miombo woodlands and geoxylic grasslands of the Cusseque area, south-central Angola. Check List 13: 2030. https://doi.org/10.15560/13.1.2030
- Sankaran M, Hanan NP, Scholes RJ, Ratnam J, Augustine DJ, Cade BS, Gignoux J, Higgins SI, Le Roux X, Ludwig F, Ardo J, Banyikwa F, Bronn A, Bucini G, Caylor KK, Coughenour MB, Diouf A, Ekaya W, Feral CJ, February EC, Frost PGH, Hiernaux P, Hrabar H, Metzger KL, Prins HHT, Ringrose S, Sea W, Tews J, Worden J, Zambatis N (2005) Determinants of woody cover in African savannas. Nature 438: 846–849. https://doi.org/10.1038/nature04070
- Sautter G (1966) De l’Atlantique au fleuve Congo, 2: Une géographie du sous-peuplement. République du Congo. République Gabonaise. De Gruyter. https://doi.org/10.1515/9783111584225
- Savadogo P, Tiveau D, Sawadogo L, Tigabu M (2008) Herbaceous species responses to long-term effects of prescribed fire, grazing and selective tree cutting in the savanna-woodlands of West Africa. Perspectives in Plant Ecology, Evolution and Systematics 10: 179–195. https://doi.org/10.1016/j.ppees.2008.03.002
- Schwartz D (1988a) Some podzols on Batéké sands and their origins, People’s Republic of Congo. Geoderma 43: 229–247. https://doi.org/10.1016/0016-7061(88)90045-6
- Schwartz D (1988b) Histoire d’un paysage: le Lousseke. Paléoéquaternaires et podzolisation sur sables Batéké (quarante derniers millénaires, région de Brazzaville, R.P. du Congo). ORSTOM, Paris. https://www.documentation.ird.fr/hor/fdi:24744 [accessed 12.04.2022]
- Schwartz D (1990) La couverture pédologique de l’Afrique centrale atlantique. In: Lanfranchi R, Schwartz D (Eds) Paysages quaternaires de l’Afrique centrale atlantique. ORSTOM, Paris, 52–59. https://www.documentation.ird.fr/hor/fdi:34768 [accessed 12.04.2022]
- Schwartz D, Lanfranchi R (1991) Les paysages de l’Afrique Centrale pendant le quarternaire. In: Lanfranchi R, Clist B (Eds) Aux origines de l’Afrique centrale. Centre Culturel Français d’Afrique Centrale; Centre International des Civilisations Bantu, Libreville, Gabon, 41–45.
- Schwartz D, Elenga H, Lanfranchi R, Marriotti A, Vincens A (1995) Les savanes du Congo: une végétation de l’holocène supérieur. In: Second Symposium on African Palynology, Orléans. Publications Occasionnelles, CIFEG: 99–108. https://www.documentation.ird.fr/hor/fdi:010004090 [accessed 12.04.2022]
- Schwartz D, Elenga H, Vincens A, Bertaux J, Mariotti A, Achoundong G, Alexandre A, Belingard C, Girardin C, Guillet B, Maley J, Namur C, Reynaud-Farrera I, Youta Happi J (2000) Origine et évolution des savanes des marges forestières en Afrique centrale Atlantique (Cameroun, Gabon, Congo) : approches aux échelles millénaires et séculaires. In: Servant M, Servant-Vildary S (Eds) Dynamique à Long Terme des Écosystèmes Forestiers Intertropicaux. IRD, UNESCO, Paris, 325–338. https://www.documentation.ird.fr/hor/fdi:010023928 [accessed 12.04.2022]
- Seranne M, Bruguier O, Moussavou M (2008) U-Pb single zircon grain dating of Present fluvial and Cenozoic aeolian sediments from Gabon: consequences on sediment provenance, reworking, and erosion processes on the equatorial West African margin. Bulletin de la Société Géologique de France 179: 29–40. https://doi.org/10.2113/gssgfbull.179.1.29
- Siebert F, Dreber N (2019) Forb ecology research in dry African savannas: knowledge, gaps, and future perspectives. Ecology and Evolution 9: 7875–7891. https://doi.org/10.1002/ece3.5307
- Silveira FAO, Ordóñez‐Parra CA, Moura LC, Schmidt IB, Andersen AN, Bond W, Buisson E, Durigan G, Fidelis A, Oliveira RS, Parr C, Rowland L, Veldman JW, Pennington RT (2021) Biome Awareness Disparity is BAD for tropical ecosystem conservation and restoration. Journal of Applied Ecology: 1365-2664.14060. https://doi.org/10.1111/1365-2664.14060
- Solofondranohatra CL, Vorontsova MS, Hempson GP, Hackel J, Cable S, Vololoniaina J, Lehmann CER (2020) Fire and grazing determined grasslands of central Madagascar represent ancient assemblages. Proceedings of the Royal Society B: Biological Sciences 287: 20200598. https://doi.org/10.1098/rspb.2020.0598
- Soret M (1973) Les Téké de l’Est. Essai sur l’Adaptation d’une Population à son Milieu. Université de Lille, France.
- Sosef MSM, Wieringa JJ, Jongkind CCH, Achoundong G, Azizet Issembe Y, Bedigian D, van den Berg RG, Breteler FJ, Cheek M, Degreef J, Faden RB, Goldblatt D, van der Maesen LJG, Ngok Banak L, Niangadouma R, Nzabi T, Nziengui B, Rogers ZS, Stévart T, van Valkenburg JLCH, Walters GHMJ, de Wilde JJFE (2006) Check-list des plantes vasculaires du Gabon. Scripta Botanica Belgica 35. National Botanic Garden of Belgium, Meise.
- Sosef MSM, Dauby G, Blach-Overgaard A, van der Burgt X, Catarino L, Damen T, Deblauwe V, Dessein S, Dransfield J, Droissart V, Duarte MC, Engledow H, Fadeur G, Figueira R, Gereau RE, Hardy OJ, Harris DJ, de Heij J, Janssens S, Klomberg Y, Ley AC, Mackinder BA, Meerts P, van de Poel JL, Sonké B, Stévart T, Stoffelen P, Svenning J-C, Sepulchre P, Zaiss R, Wieringa JJ, Couvreur TLP (2017) Exploring the floristic diversity of tropical Africa. BMC Biology 15: 15. https://doi.org/10.1186/s12915-017-0356-8
- Stohlgren TJ, Barnett DT, Simonson SE (2002) Beyond NAWMA-The North American Weed Management Association Mapping Standards.
- Stokes S, Thomas DSG, Washington R (1997) Multiple episodes of aridity in southern Africa since the last interglacial period. Nature 388: 154–158. https://doi.org/10.1038/40596
- Stone RD (2014) The species-rich, paleotropical genus Memecylon (Melastomataceae): Molecular phylogenetics and revised infrageneric classification of the African species. Taxon 63: 539–561. https://doi.org/10.12705/633.10
- Stone RD, Walters G, Ghogue J-P (2006) Memecylon batekeanum, a new species from southeastern Gabon, and a note on the circumscription of Memecylon sect. Mouririoidea (Melastomataceae, subfamily Memecyloideae). Novon 16: 276–280. https://doi.org/10.3417/1055-3177(2006)16[276:MBANSF]2.0.CO;2
- Swift J (1996) A brief history of desertification. In: Leach M, Mearns R (Eds) The Lie of the Land: Challenging Received Wisdom on the African Environment. James Currey, Oxford, England, 73–90.
- Systat Software Inc. (2007) SYSTAT® 12 Statistics_I_II_III_IV. Version 12.0. Systat Software, Inc., San Jose, California.
- Thiers B (continuously updated) Index Herbariorum: A global directory of public herbaria and associated staff. New York Botanical Garden’s Virtual Herbarium. http://sweetgum.nybg.org/ih/ [accessed 21.04.2022]
- Thomas DSG, Shaw PA (1991) The Kalahari Environment. Cambridge University Press, Cambridge. https://archive.org/details/kalaharienvironm0000thom [accessed 12.04.2022]
- Ukizintambara T, White L, Abernethy K, Thébaud C (2007) Gallery forests versus bosquets: conservation of natural fragments at Lopé National Park in central Gabon. African Journal of Ecology 45: 476–482. https://doi.org/10.1111/j.1365-2028.2007.00757.x
- University of Maryland (2006) FireMapper Database. https://modis-fire.umd.edu/ [accessed 21.01.2022]
- Uys RG, Bond WJ, Everson TM (2004) The effect of different fire regimes on plant diversity in southern African grasslands. Biological Conservation 118: 489–499. https://doi.org/10.1016/j.biocon.2003.09.024
- Vansina J (1990) Paths in the Rainforest: Toward a History of Political Tradition in Equatorial Africa. James Currey, London. https://archive.org/details/pathsinrainfores0000vans [accessed 12.04.2022]
- Veldman JW, Buisson E, Durigan G, Fernandes GW, Le Stradic S, Mahy G, Negreiros D, Overbeck GE, Veldman RG, Zaloumis NP, Putz FE, Bond WJ (2015a) Toward an old-growth concept for grasslands, savannas, and woodlands. Frontiers in Ecology and the Environment 13: 154–162. https://doi.org/10.1890/140270
- Veldman JW, Overbeck GE, Negreiros D, Mahy G, Le Stradic S, Fernandes GW, Durigan G, Buisson E, Putz FE, Bond WJ (2015b) Where tree planting and forest expansion are bad for biodiversity and ecosystem services. BioScience 65: 1011–1018. https://doi.org/10.1093/biosci/biv118
- Veldman JW, Overbeck GE, Negreiros D, Mahy G, Le Stradic S, Fernandes GW, Durigan G, Buisson E, Putz FE, Bond WJ (2015c) Tyranny of trees in grassy biomes. Science 347: 484–485. https://doi.org/10.1126/science.347.6221.484-c
- Vincens A, Schwartz D, Elenga H, Reynaud-Farrera I, Alexandre A, Bertaux J, Mariotti A, Martin L, Meunier J-D, Nguetsop F, Servant M, Servant-Vildary S, Wirrmann D (1999) Forest response to climate changes in Atlantic Equatorial Africa during the last 4000 years BP and inheritance on the modern landscapes. Journal of Biogeography 26: 879–885. https://doi.org/10.1046/j.1365-2699.1999.00333.x
- Walters G (2010) the Land Chief’s embers: ethnobotany of Batéké fire regimes, savanna vegetation and resource use in Gabon. PhD thesis, University College of London, UK. https://discovery.ucl.ac.uk/id/eprint/20195/ [accessed 12.04.2022]
- Walters G (2012) Customary fire regimes and vegetation structure in Gabon’s Bateke Plateaux. Human Ecology 40: 943–955. https://doi.org/10.1007/s10745-012-9536-x
- Walters G (2015) Changing fire governance in Gabon’s Plateaux Bateke savanna landscape. Conservation and Society 13: 275–286. https://www.jstor.org/stable/26393206 [accessed 12.04.2022]
- Walters G, Bradley A, Niangadouma R (2006) Floristics of Gabon’s Batéké Plateaux: Guineo-Congolian plants on Kalahari Sands. In: Gazanfar SA, Beentje HJ (Eds) Taxonomy and Ecology of African Plants, their Conservation and Sustainable Use. Royal Botanic Gardens Kew, Richmond, 259–266.
- Walters G, Parmentier I, Stévart T (2012) Diversity and conservation value of Gabon’s savanna and inselberg open vegetation: an initial gap analysis. Plant Ecology and Evolution 145: 46–54. https://doi.org/10.5091/plecevo.2012.606
- Walters G, Touladjan S, Makouka L (2014) Integrating cultural and conservation contexts of hunting: the case of the Plateaux Batéké savannas of Gabon. African Study Monographs 35(2): 99–128.
- Walters G, Schleicher J, Hymas O, Coad L (2015) Evolving hunting practices in Gabon: lessons for community-based conservation interventions. Ecology and Society 20: art31. https://doi.org/10.5751/ES-08047-200431
- Walters G, Sayer J, Boedhihartono AK, Endamana D, Angu Angu K (2021) Integrating landscape ecology into landscape practice in Central African Rainforests. Landscape Ecology 36: 2427–2441. https://doi.org/10.1007/s10980-021-01237-3
- Walters G, Touladjan S, Christy P, Assaly RB, Bout N, Pearson L (2010) Interdisciplinary studies: understanding fire management using social an biological data. In: Vololoniaina HJ, Razafimandimbison SG, De Block P (Eds) XIXth AETFAT Congress, Madagascar, 25–30 April 2010, Abstracts. Scripta Botanica Belgica 46, National Botanic Garden of Belgium, Meise, 498.
- White F (1976) The underground forests of Africa: a preliminary review. Gardens’ bulletin, Straits Settlements 29: 57–71.
- White F (1979) The Guineo-Congolian Region and its relationships to other phytochoria. Bulletin du Jardin botanique national de Belgique / Bulletin van de National Plantentuin van België 49: 11–55. https://doi.org/10.2307/3667815
- White F (1983) La Végétation de l’Afrique: Mémoire Accompagnant la Carte de Végétation de l’Afrique. ORSTOM, Paris. https://www.documentation.ird.fr/hor/fdi:24837 [accessed 12.04.2022]
- Wieringa JJ, Sosef MSM (2011) The applicability of Relative Floristic Resemblance to evaluate the conservation value of protected areas. Plant Ecology and Evolution 144: 242–248. https://doi.org/10.5091/plecevo.2011.588
- van Wilgen BW, Govender N, Biggs HC (2007) The contribution of fire research to fire management: a critical review of a long-term experiment in the Kruger National Park, South Africa. International Journal of Wildland Fire 16: 519. https://doi.org/10.1071/WF06115
Supplementary materials
Checklist of the Parc National des Plateaux Batéké and a 25 km buffer zone
List of forb and geoxylic suffrutices species in the burn experiment
Plot design for the fire study. Figure adapted from Stohlgren et al. (2002)